


Abstract
Although not fully understood, sleep is accepted as a vital and organized sequence of events that follows a regular cyclic program each night to ensure the human body can perform at its optimum. A lack of sleep, or sleep deprivation (SD), is a widespread phenomenon that can induce adverse changes in cognitive performance. This review focused on the biological explanation as well as the research investigating the numerous effects that SD can have on cognition. A reduction in sleep does not occur independently of the effects on memory, attention, alertness, judgment, decision-making, and overall cognitive abilities in the brain, resulting in decreased function and impaired cognitive performance.
Sleep deprivation (SD) can be divided in partial and total SD. Partial SD refers to a night of reduced or interrupted sleep, and total SD to no sleep for at least one night during the normal sleep/wake cycle. Total SD is frequently studied, which could be due to the easier monitoring of brain activity as longer periods of SD cause increased impaired cognitive functioning. Studies show that consistently restricting sleep overtime is more harmful in comparison to a single night of total SD.1
When sleeping, an individual experiences 3 NREM (non-rapid eye movement) and one REM (rapid eye movement) sleep phases. REM SD appears to have a notable effect on exciting neurons, which is vital for assessing possible danger as well as processing reactions to stimuli associated with threats. The NREM sleep deprivation reduces the normal release of specific neurotransmitters, which can affect the ability of the receptors to refresh and restore sensitivity.2 Without these stages of sleep, the result is reduced cognition.
Two processes that affect the amount of sleep obtained are the circadian and homeostatic processes. The circadian process theory proposes a regulating internal circadian clock, which causes the onset and offset of sleep incidents. This facilitates the control wake and sleep cycles as well as the secretion of different hormones. The homeostatic process depends on the wakeful and sleep periods, and the sleep requirements increase with inadequate sleeping hours. As SD increases, the homeostatic functions of the brain become increasingly impaired. The brain attempts to counter this by secreting hormones in a wave-like manner to prompt wakefulness or sleep.3 The interaction of the 2 processes regulates the sleep/wake cycle.
The SD effects on judgement and decision- making
Individuals with SD loose the functional connectivity between the amygdala and the medial prefrontal cortex (mPFC), a region known for exhibiting strong inhibitory projections to the amygdala.4 In addition, a higher connective relationship occurs in the autonomic areas of the locus coeruleus and the amygdala (Figure 1).
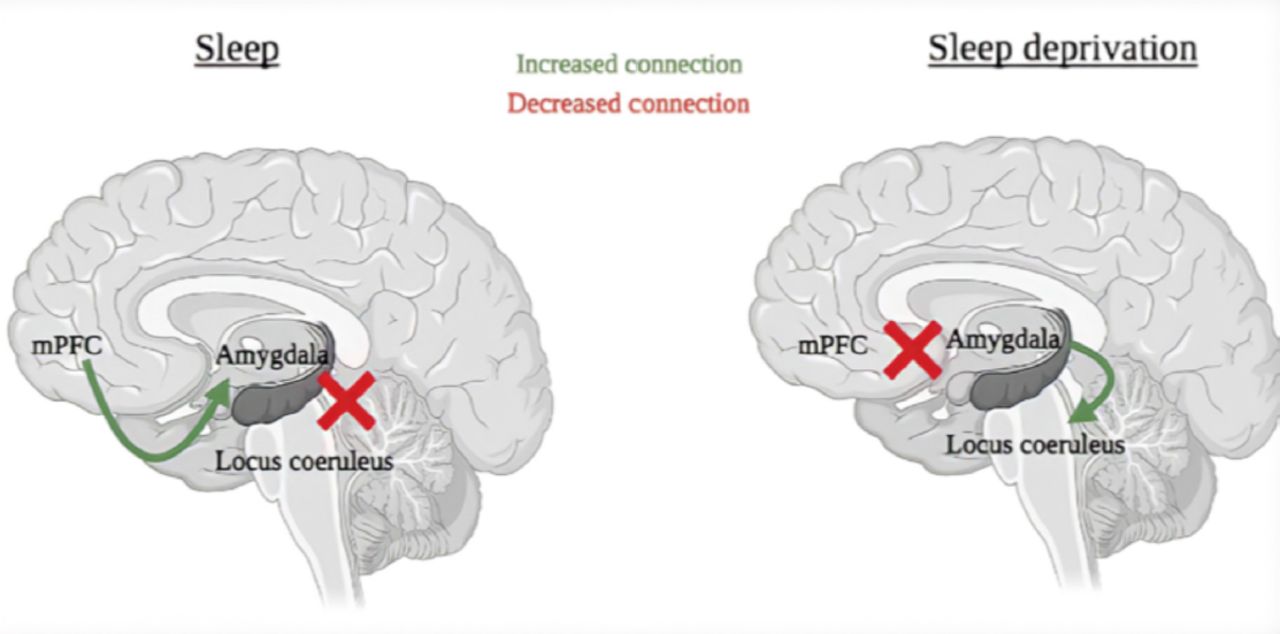
- Comparison of the connections between the mPFC and amygdala, as well as the amygdala and the locus coeruleus, during normal sleep vs SD. SD - Sleep deprivation, mPFC - medial prefrontal cortex
With a lack of sleep, an increased amygdala hyperlimbic reaction occurs, resulting in stimuli with negative emotional connotations.5 This varying level of amygdala activity is linked to a loss of mPFC functional connectivity when sleep deprived, suggesting a decrease in prefrontal lobe inhibition signals. SD results in missing a corrective brain reset reactivity due to the dysfunctional integrity of the mPFC-amygdala circuit, causing inappropriate behavioral responses, such as making rational decisions and social judgements.6 A study examining moral judgement in individuals with SD reported that SD causes longer response latencies, suggesting a higher level of difficulty to decide a suitable course of action, as SD impairs the ability to integrate cognition and emotion to pass moral judgment.7
However, the study used a debatable single assessment procedure of moral judgment, which can limit the generalizability of the results.
The EEG scans propose a noticeable function for the medial prefrontal cortex in the development of ethical judgments and decision-making. Unique and significant changes were found in the mPFC in short- term SD participants, with a further demonstration of how the mPFC attempts to compensate for cognitive failing during SD. Movement in this area is also significantly reduced following SD.8 In contrast, a study claimed that short-term SD does not selectively affect prefrontal functioning. However, all the tests carried out in this study were derived from a neuropsychological battery test created for clinical purposes, mostly to examine brain damage. The tests could have a ceiling effect, not influenced by short-term SD, and the participants were all university students.9
Space Shuttle Challenger Accident acknowledged the role of poor judgment and human error related to early hour shift work and sleep loss, stating that the decision to launch should have been based on engineering judgments (V, Washington, D.C, 1986). Despite these comments, consideration should be given to other factors which may have influenced proceedings related to successful exchange communication and information.
The SD effects on memory
Sleep deprivation appears to disrupt memory consolidation in the hippocampus through long-term potentiation (LTP). The NMDA receptor is required for the consolidation phase of memory, as it allows memory to advance from an unstable to a more permanent form, which is disrupted in SD.10 Gais et al11 found that the NDMA receptors allow the expression of LTP through an increased Ca2+ influx. SD can impair the full activation of the glutamate receptor by altering the surface and receptor sub-unit composition expression, with a reduced Ca2+ influx. SD rats showed a reduction in tthe NMDA/AMPA ratio of the hippocampal cells, following hippocampal synapse stimulation. Although there were no alterations in the glutamate or channel sensitivity, it was observed that SD changes the NMDAR surface expression after a crosslinking assay indicated that the NR2A and NR1 components of the NMDA receptors remained in the cytoplasm after SD.12
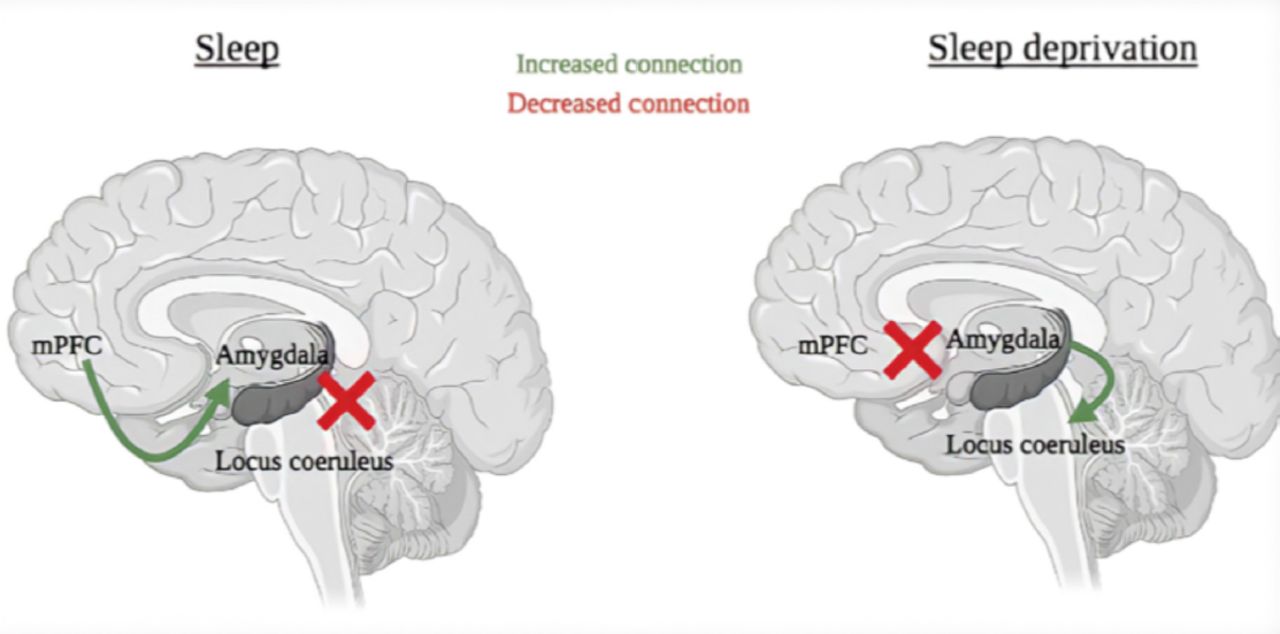
Figure 2A indicates how sleep deprivation alters glutamatergic signaling through modifications in AMPA and NDMA receptor structure. This disturbance in the receptor function can reduce the molecular signaling cascades due to an attenuated calcium influx, resulting in fewer permanent memories being consolidated in the brain.13

- An overview of signaling pathways in the hippocampus following sleep deprivation. The dashed arrows demonstrate a reduction of a signaling pathway. (A) Altered glutamatergic signaling. disrupted cAMP signaling. (C) Down regulated mTOR signaling. Figure adapted from Prince and Abel14
The hippocampal activation of the Protein Kinase activity (PKA) and Cyclic adenosine monophosphate (cAMP) signaling plays a vital role in memory strengthening. Phosphodiesterase 4A (PDE4A) is an enzyme responsible for the degradation of cAMP in the hippocampus, causing a reduction in the protein kinsase A (PKA) activity. The SD follows with an enhanced activity of the PDE4 and PDE4A5 protein. Blocking the PDE4 signaling results in increased LTP, and reduces SD deficits in the hippocampus-dependent memory consolidation. This rescue by PDE4 inhibition indicates that this molecular disruption by SD produces the functional deficits in plasticity and behavior.14 A study investigated whether the SD increased the PDE4 levels in the hippocampus of mice. It was found that the PDE4-specific cAMP breakdown increased in SD mice, however the non-PDE4 levels were unaffected.15 The increase in the PDE4A by SD disrupts cAMP intracellular signaling (Figure 2B).
The SD also down-regulates the mammalian target of rapamycin (mTOR) signaling, a translational regulatory protein required for memory consolidation (Figure 2C). This implies that protein translation is necessary for plasticity and long-standing memory development. However, more studies are required to explore the changes in the gene transcription as well as protein synthesis following SD.16
A low level of SD is sufficient to activate the unfolded protein response (UPR), which results in a reduction of protein synthesis.17 The initiation of plastic modifications during an aroused state can increase the need for protein synthesis; however, after wake surpasses its physiological duration in SD, protein synthesis is compromised. The decreased energy consumption by the synaptic transmission in SD indicates that sleep in slow waves may signify an interval for the brain cells to conduct many roles, including membrane recycling, restoring glutamate vesicles, protein translation, restoring calcium in the presynaptic stores, and mitochondrial rest.18
An increase in SD can result in a decrease in these functions which can negatively affect memory consolidation.19 In particular, the inhibition of the RbAp48 protein in young mice caused hippocampus-dependent memory impairment. FMRI studies showed dysfunction in the hippocampal formation, with a selective reduction in histone acetylation, leading to memory loss.20 Although we can conclude that SD can cause decreased protein synthesis of certain proteins such as RbAp48, which can result in reduced memory, this study did not observe the effects of SD in the mice to be able to conclude whether sleep is the main cause of reduced memory.
The temporal lobe serves the purpose of memory encoding and language processing. A lack of sleep can result in the incapacity of the brain to process neural signals at optimal quantities causing incoherent speech.21 Studies show that SD can disrupt the role of NREM sleep in downregulating the histamine, serotonin and norepinephrine neurotransmitters. This means the receptors lack the capacity to “rest” and restore sensitivity. In turn, the norepinephrine, serotonin and histamine cannot function optimally and are not produced at natural levels, leading to an impairment in the storage of memory and day- to-day speech.22 The SD also results in a lack of the enzymes that repair brain cell damage caused by free radicals, again affecting memory and speech.23 This can worsen for people with long term SD as their neurons start to degenerate due to the relentless brain activity.24 However, Cirelli et al25 found opposing results with no evidence of brain cell degeneration after long-term SD in rats, indicating more research is required the determine the cause of the neuron degeneration.
The SD effects on attention and alertness
In the sleep state, there is an equal inhibitory action of the task-related default mode network (DMN) activity and the front parietal network (FPN) activity. This is supported by the increased arousal from the thalamus. This equal inhibition means there is consistent attentional performance in the brain.26 The DMN is the collection of brain areas that activate and deactivate depending on an individual’s external tasks, while the FPN are the networks and brain regions linked to attention (Figure 3A). In a sleep-deprived state, there is an imbalanced inhibition between the task-related DMN and FPM activity, and inconsistent increasing arousal activity that influences the thalamic activity.27 This results in an irregular disturbance of the DMN activity and a reduced FPN activity during external tasks. Suppressing the DMN is vital to allow appropriate brain networks to achieve successful behavior towards tasks and goals, and without its suppression, an individual is unable to maintain attention to specific tasks. Once FPN activity and DMN action are altered, attention and concentration tasks will be unpredictable (Figure 3B). After sleep is obtained, the arousal input from the thalamus will stabilize and allow the equal activity of the DMN and FPN to be re-established.28

- Areas affected by SD (A) Areas of the brain affected by sleep deprivation, affecting alertness, memory and attentional performance. (B) In a SD state, there is unstable inhibition concerning task-related DMN and FPN activity, as well as inconsistent increasing arousal influencing activity in the thalamus. This leads to irregular signals of DMN activity and reduced FPN activity during tasks. This can lead to weakened attentiveness and working-memory functioning, becoming better with greater thalamic activity and poorer with reduced thalamic activity. Figure adapted from Krause et al54
According to a study conducted on objective attention, SD resulted in slower and imprecise picture sorting in addition to a lower memory detection for the pictures. Activation in the parahippocampal place area (PPA) and the fronto-parietal cortex in response to task-related activity was reduced in SD. This reduction in effective integration and functional connectivity of the sub parts of a task affects the ability to perform a task successfully.29 Visual tasks given to participants showed that the difficulty of the tasks was related to parietal cortex activation, and inactivation of the insular cortices, visual cortices, and the cingulate gyrus. This pattern of activation and deactivation was considerably lower in the SD participants than the group with complete sleep. These combined factors can cause impairment in the attentional networks essential for accurate attention performance and can lead to higher vulnerability to risks and accidents in routine life.30 The PET scans indicated decreased brain activity primarily in the thalamus and the prefrontal cortex, which are both associated with attention and alertness.
This could be explained by the significant decrease in the relative regional glucose observed predominantly in the thalamus and prefrontal and posterior parietal cortices, indicating that alertness and attention is indirectly impacted by increased fatigue due to sleep loss.31 However, in contrast, Kuhn et al32 found an increase in the glucose levels in SD individuals, indicating that further research may be required to confirm the effect of the glucose levels on attention and alertness in SD individuals. In a SD state, there is unstable inhibition of task-related DMN and FPN activity, as well as an inconsistent increasing arousal influencing activity in the thalamus. This causes irregular signals of DMN activity and reduced FPN activity during tasks. This can cause a loss of attentiveness and working-memory functioning, improving with greater thalamic activity and less with reduced thalamic activity.
The EEG displays in rats indicate that SD results in more cortical neurons showing periods of silence, identical to the OFF periods linked with the slow oscillations of sleep.33 These OFF periods during wake may result from the heightened neural bistability as a result of the increased inclination to hyperpolarization. This hyperpolarization causes the membrane potential to become more negative in neurons, and may be an outcome of synaptic surplus due to powerful wake plasticity. This can lead to an unevenness of the supply and demand energy, signaled by an increase in extracellular adenosine.34
It is unclear whether OFF stages are able to undergo a few of the healing roles of sleep, namely synaptic homeostasis. Nevertheless, the existence of OFF periods during wake periods offer interesting explanations. For example, the dissociation between attentiveness and specific cognitive functions during SD can be explained by the incidence of local sleep in arousal subcortical promoting systems, and at additional periods in precise cortical regions.35 In addition, if local sleep-in wake occurred in the brainstem and hypothalamic nerve cells that apply control in arousal, it could clarify the reasons for the augmented global impairments and sleepiness in attention following SD, especially for less demanding uninteresting activities. Testing overall performance in attention and alertness in SD subjects is very unstable, ranging from normal levels to dangerous mistakes.36
The synaptic homeostasis hypothesis
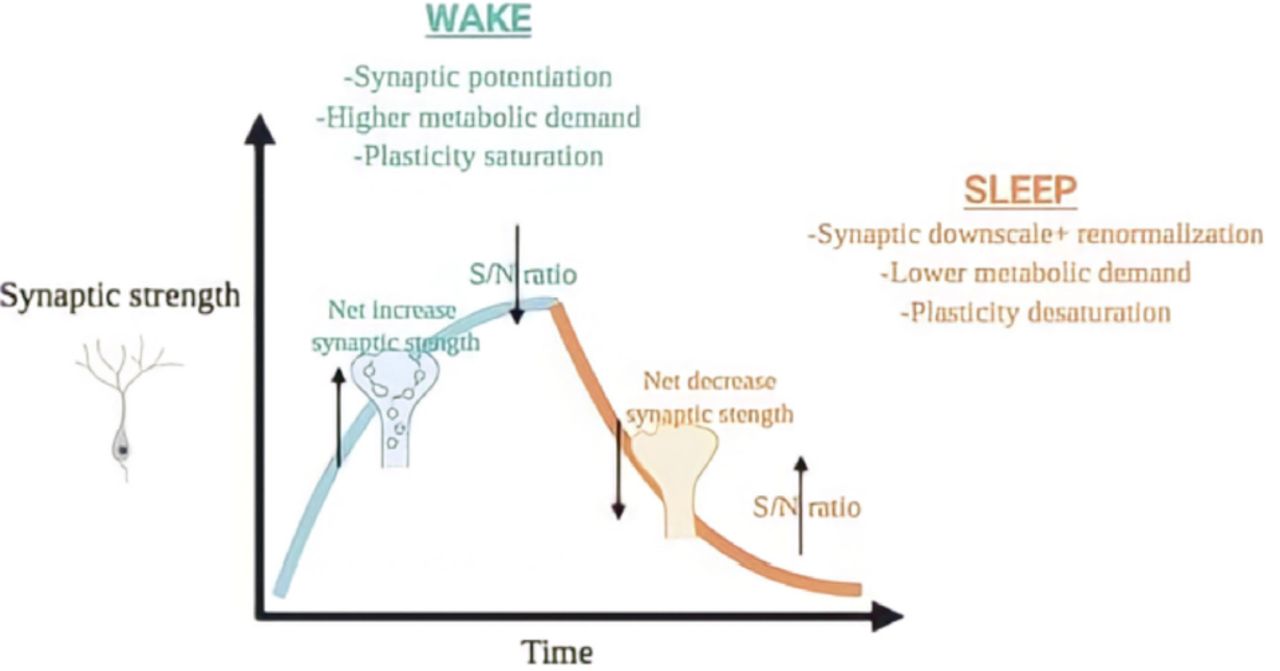
Shown in Figure 4, the synaptic homeostasis hypothesis (SHY) proposes that during the slow wave stage of NREM sleep, synapses are decreased throughout the brain to counteract the net strengthening of network synapses during a waking experience, such as learning.37 During wakefulness, absorbing the surrounding atmosphere necessitates solidifying networks within the brain.38 This requires an increase of the cellular processes for energy, learning consolidation, and a reduction of the signal-to-noise ratios.39 Spontaneous activity, which is any neural activity not driven by a task, can renormalize the synaptic strength and restore cellular homeostasis during sleep.

- The synaptic homeostasis hypothesis. Throughout wakefulness, cortical synapses potentiate relative to activity, giving a net rise in synaptic power and a reduction is signal/noise ratio. This synaptic potentiation is linked with an escalation in slow-wave activity (SWA) in NREM sleep, where synaptic renormalization and downscaling occurs, with an increase in signal/noise ratio. This mechanism can permit additional synaptic plasticity to happen the resulting day alongside avoiding the metabolic downsides linked with excitability and extreme potentiation through wakefulness. Figure adapted from Rantamäki and Kohtala.37
The strength reduction of synapses can explain the benefits of sleep on memory acquisition and consolidation, as energy is saved when counteracting the network effects of synaptic excitation and increased neuronal activity following wake periods.40 Differing results can follow an amplification in the synaptic strength, comprising faster energy utilization, increased need for the distribution of cellular components to synapses which causes stress to cells, and modifications in the glia support cells.41 This increased synaptic power can also weaken the selectivity of neuronal reactions and consolidates the skill to absorb information.
Without normalizing the synaptic power, SD will increase the weight of plasticity on the nerve cells and fail to re-establish neuronal selectivity and learning abilities, which does not allow enhancing of the signal-to-noise ratios. As SD continues, the lack of sleep will diminish the brain’s ability to re-establish cellular and synaptic homeostasis challenged by plastic changes which happens during normal wake, resulting in a consolidation and integration of memories.42,43
Synaptic renormalization displays the many benefits of sleep, through the desaturation of the synaptic networks, the enhancement in energy provision, and the decrease in cellular pressure causing an improved learning experience and retention of memories.44 For instance, the coding of new images is compromised after a night of slight SD, which reduces the slow wave activity, exclusive of lessening total SD.45 However, Gais et al11 concluded through fMRI studies that declarative memory is not affected with long term SD, and Voderholzer et al46 that long term SD does not affect long term declarative memory in adolescents.
Similarly, episodic memory retention can be significantly reduced if a training task is carried out after SD, regardless of no alterations in reaction time during the training, which suggests a reduction in coding ability because of SD.6 Opposing this, an increased encrypting ability of pictures, word pairs, and lists were observed following a short sleep in which slow oscillations were boosted by transcranial and sham stimulation. This was possibly due to the down-scaling of synaptic networks in the hippocampus, that were potentiated towards saturation during the previous period of wakefulness.47
The brain’s glymphatic system
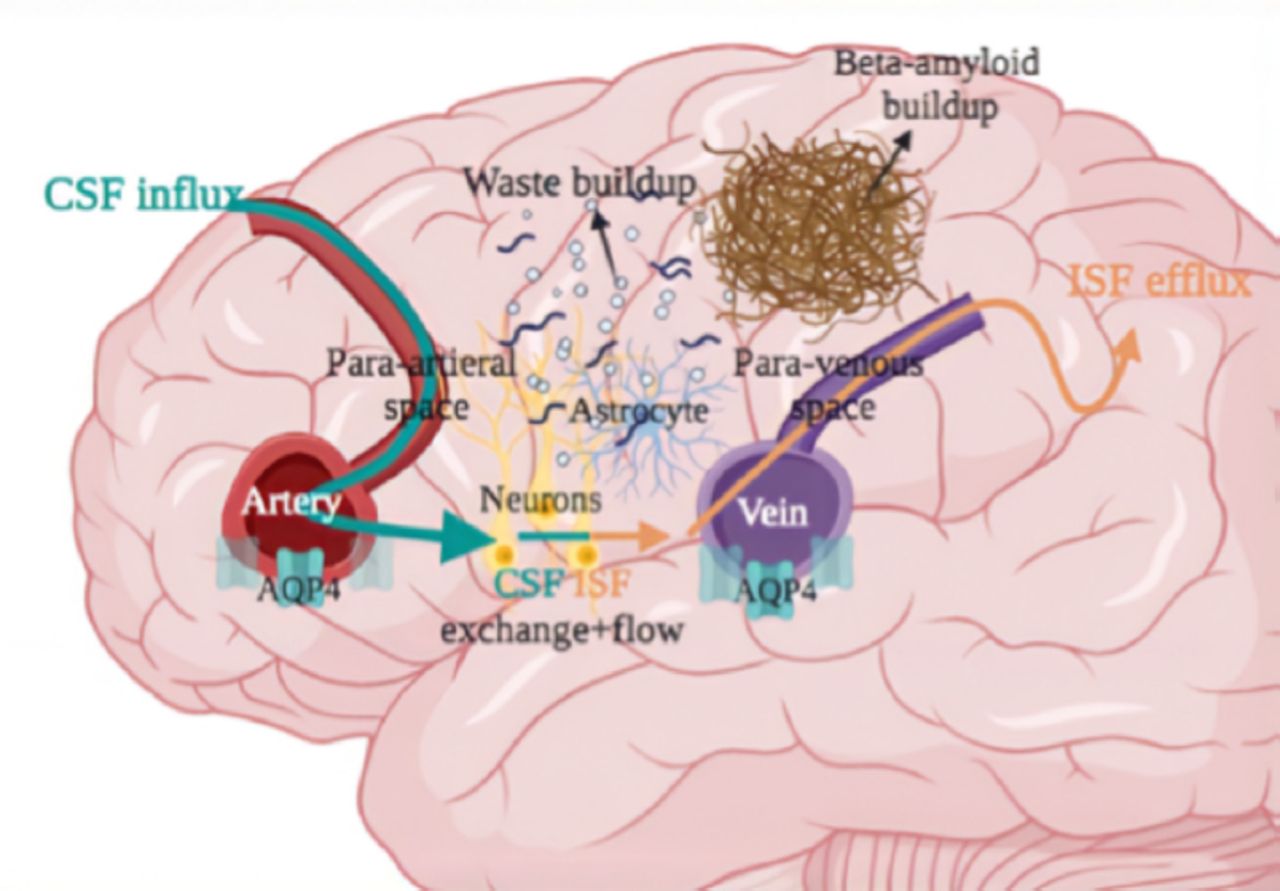
The glymphatic system consists of a complex of vessels that remove waste from the central nervous system, typically in the sleep state. This happens when the cerebrospinal fluid (CSF) moves into the para-arterial region, followed by an interstitial area across the aquaporin 4 (APQ4) water channel route. Due to rhythmic pulsations during the stages of sleep, an interchange arises between the interstitial fluid (ISF) as well as the CSF. Unused products are relocated from the arteries to the veins, which travel across to the paravenous region.48
The reduced expression of the water channel pathway, AQP4 can increase the probability of additional secretion of a peptide called beta-amyloid. The sleep and awake states have different glymphatic system roles. During the sleep state, the CSF flows at increased levels, and the interstitial space increases by 60%, which can successfully clear toxic cellular molecules. Following a sleep-state, the beta-amyloid levels can be reduced by the CSF. Beta amyloid is a primary reason for many neurodegenerative diseases such as Dementia and Alzheimer’s disease. This shows that sleep deprivation can diminish the active process of the glymphatic system, leading to toxin build-up which can negatively affect the cognitive performance, motor functions and behavioral patterns.49 (Figure 5).

- Impaired glymphatic system in the brain after sleep deprivation. Lower AQP4 expression, altered glymphatic clearance, toxic waste buildup, and higher beta-amyloid levels occur, leading to a dysfunction in cognitive performance.
The reduction of the glymphatic activity also results in a dysregulation of the apolipoprotein E circulation in the brain, dysregulation of the glucose metabolism, insulin resistance, resulting in cognitive decline. Overtime, these metabolite accumulation and dysregulation can cause severe brain complications.50 PET scans with tracers were used to study the amount of amyloid-beta in mice through standard sleep and sleep deprivation. A one-night comparison showed that there was a significant increase in the beta-amyloid levels in the thalamus and the hippocampus of the mice, demonstrating in vivo evidence of the effects of sleep deprivation on recognized neurodegenerative processes.51
Despite these findings, the PET scan technique could not distinguish between the soluble and insoluble beta-amyloid, which could affect the findings as soluble, rather than insoluble amyloid-beta, is more predictive of neurodegenerative disorders such as Alzheimer’s disease. Another study with older participants found an inverse relationship between the beta-amyloid and performance in several cognitive areas, with an increase in the beta-amyloid. This can be indicative of a deficient glymphatic system in individuals with SD, which cause a lower performance in global cognition, verbal fluency, confrontation naming, and verbal learning.52
However, SD was not studied in the participants and so more research is required to conclude whether the increase of beta amyloid is indeed due to an impaired glymphatic system, specifically due to sleep loss. Opposing evidence in the Melzer et al study53 fails to find a connection between the beta-amyloid deposition and a decline in cognitive performance, suggesting there are other factors which could be the primary driver of the cognitive impairment.
Conclusion
Sleep is undoubtedly a vital component in a healthy lifestyle. To maintain our bodies in a consistent wake and sleep cycle, the brain continually secretes neurological hormones. The adverse consequences of SD are evident on overall behaviour and cognitive performance. Neurological pathways slow down, causing a reduced reaction time and mental state. Systems in the body enters a state of life-support as a means of coping until the brain can be re-stimulated. Fluctuations in the thalamic activity, synaptic renormalization, glymphatic system roles, DMN activity, amygdala activity and hippocampal activity can cause unequal stimulation in the brain, which results in irregular activities in the brain to manage SD. As a result, an impairment in attentiveness, working memory, consolidation of memories, alertness, judgement, decision-making, and many other diminished cognitive performances will follow. After having the required quantity of sleep for a specific person, the brain will be at an advantage with higher reaction times and intellectual activity. Additional research is required to provide evidence of the validity of the exact mechanisms and subsequent effects of SD, which can be achieved with more resources, study and time. Despite this, current evidence provide proof that regardless of health, receiving inadequate sleep daily or for multiple days causes the body’s systems to gradually decline. The human mind and body need sufficient NREM and REM stages of the sleep phases, to have enough sleep and maximize brain function by keeping cognitive performance at an optimum.
Acknowledgment
The authors acknowledge Ms. Lalona Khan for drawing and producing images.
Footnotes
Disclosure. The authors declare no conflicting interests, support or funding from any drug company.
- Copyright: © Neurosciences
Neurosciences is an Open Access journal and articles published are distributed under the terms of the Creative Commons Attribution-NonCommercial License (CC BY-NC). Readers may copy, distribute, and display the work for non-commercial purposes with the proper citation of the original work.
In this issue

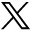


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.