


In contradiction to the crossing of all retinal fibers in the lateral-eye species, the retinal fibers of binocular vision in frontal-eye species run either ipsilaterally or cross the midline twice to terminate the ipsilateral cerebral hemisphere.1 It is hypothesized that carrying separated information, such as vision in lateral eyes, seems to create the need for functional coordination between the 2 cerebral hemispheres. This coordination appears to be mediated by fiber crossing. Shared information; however, such as binocular vision and olfactory pathway, runs ipsilaterally. In this situation, both cerebral hemispheres receive the same information; therefore, functional coordination between the 2 hemispheres via fiber crossing is mostly unnecessary.1 In another view,2 the eye-forelimb hypothesis suggests that the ipsilateral retinal fibers of binocular vision as the right fibers of the right retina carry binocular vision from the left hemifield of the right visual field and run ipsilaterally to reach the right cerebral hemisphere. The latter controls the left limbs; thus, enabling more target-oriented motor response to stimuli in the left hemispace including the left hemifield of the right visual field. According to the last hypothesis,2 if these described fibers were to cross the midline to the left cerebral hemisphere, the same fibers would have to cross the midline again to the right cerebral hemisphere to control the left limbs. This obviously would elongate the pathway and probably would add at least another synaptic link. Subsequently, this would make the motor response to stimuli in the binocular visual hemifields slower and less effective. Larsson2 noted that in the owl, binocular vision is mediated by long neural pathways with double crossing and more synaptic links, despite rapid and effective motor response to stimuli in the binocular visual hemifields.
In this present paper, we propose an alternative hypothesis, ‘ipsilaterality serves contralaterality’, for the explanation of the presence of ipsilateral retinal fibers of binocular vision. This hypothesis relies on the well-documented principle of contralaterality, which dominates the neural pathways in the CNS.1 In this concept, each cerebral hemisphere deals with the contralateral hemispace. For instance, the right cerebral hemisphere receives sensory stimuli from the left hemispace through the right halves of the 2 retinae. The binocular visual fibers; namely, the right fibers of the right retina and the left fibers of the left retina run ipsilaterally to reach the right and the left hemispheres contralateral to their binocular visual hemifields; namely, the left hemifield of the right visual field and the right hemifield of the left visual field.
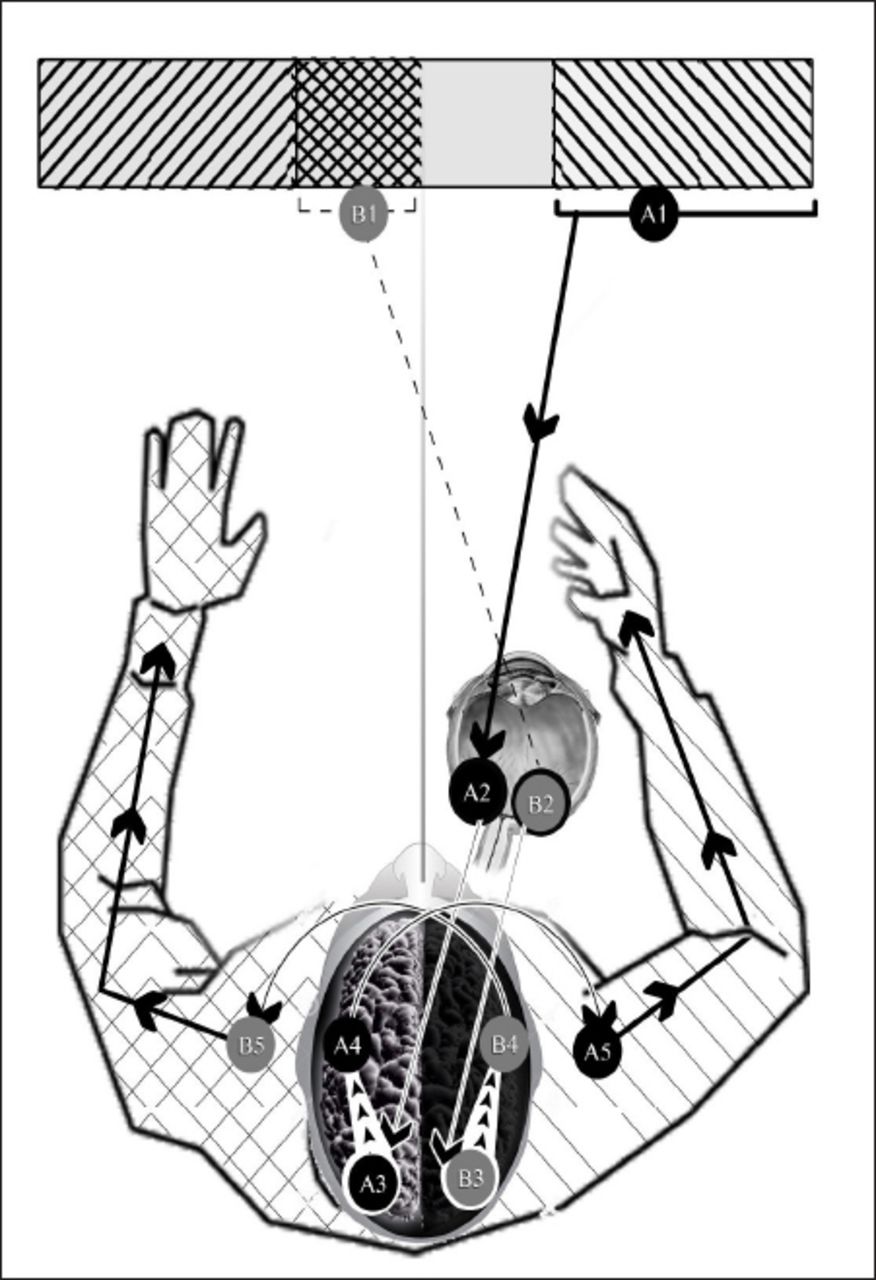
In lateral eyes, such as in birds, the 2 hemifields of the right visual field are located in the right hemispace and all the stimuli from the right visual field reach the left cerebral hemisphere after they cross the midline. In frontal-eyes with binocular vision, the left hemifield of the right visual field is located in the left hemispace and the retinal fibers of the right eyes that carry stimuli from this left hemifield of the right visual field run ipsilaterally to reach the right cerebral hemisphere, which lie contralateral to the left hemispace.
Apparently, when lateral eyes shifted to frontal-eyes during evolution (Figure 1) the left hemifield of the right visual field shifted from the right hemispace to the left hemispace while the right hemifield of the right visual field remains in the right hemispace. According to the concept of contralaterality, binocular stimuli in the left hemifield of the right visual field must shift their projection to the right cerebral hemisphere. Therefore, the retinal fibers that carry these stimuli; such as, the right fibers of the right retina must shift their projection to the right cerebral hemisphere namely run ipsilaterally. If the right fibers of the right retina were to keep their projection to the left cerebral hemisphere after shifting from lateral eyes to frontal-eyes, stimuli from the left hemifield of the right visual field, as now part of the left hemispace, would project to the left cerebral hemisphere, which are ipsilateral to the left hemispace. This would not only contradict the principle of contralaterality, but it would also render the motor response of the limbs to stimuli less effective. The right limbs that are controlled by the left cerebral hemisphere would not respond target- and side-oriented to stimuli in the left hemispace, thus jeopardizing survival chances.

Frontal eye model in humans. A1 - right hemifield of right visual field, A2 - nasal retina of right eye, A3 - visual cortex of left cerebral hemisphere, A4 - motor cortex of left cerebral hemisphere, A5 - right upper limb, B1 - left hemifield of right visual field, B2 - temporal retina of right eye, B3 - visual cortex of right cerebral hemisphere, B4 - motor cortex of right cerebral cortex, B5 - left upper limb. Visual hemifields A1 and B1 are similarly shadowed as right upper limb and left the upper limb.
Shifting of projection of stimuli in the left hemifield of the right visual field from the left cerebral hemisphere in lateral eyes to the right cerebral hemisphere in frontal-eyes, certainly follows the principle of contralaterality and most probably enables the right cerebral hemisphere to control the response of left limbs to stimuli in the left hemispace in a more effective fashion. Similar conclusions apply to the right hemifield of the left visual field and the left cerebral hemisphere. The general survival chances are subsequently improved.
In conclusion it can be argued that with the advent of binocular vision in frontal-eyes shifting; thus, overlapping of visual hemifields of either side happened. The evolved corresponding retinal fibers subsequently developed new ipsilateral cerebral projections. In this new arrangement, the ipsilateral cerebral hemispheres lie contralateral to their corresponding binocular visual hemifields. The ipsilateral retinal fibers of binocular vision; namely the temporal fibers of the 2 retina run ipsilaterally to reach cerebral hemispheres contralateral to their visual hemifields. This behavior appears to be a modification of the lateral eye model where, in conformation with other sensory pathways, all retinal fibers cross the midline. One benefit of this modification appears to enable each cerebral hemisphere to deal with visual sensory stimuli and control muscular activity in the contralateral hemispace. The limbs of either side can therefore react to sensory stimuli in the same hemispace. Thus, ipsilateral pathway serves the classical contralateral control. It should be noted that the ipsilateral retinal fibers of binocular vision follow the principle of contralaterality, whether through short uncrossed pathways in some frontal-eye species such as the human, or through long double crossed pathways in other frontal-eye species such as the owl. These ipsilateral pathways of projection can be assumed to serve the old concept of contralateral cerebral control, and can be interpreted as a step further raising the scores of survival chances and improving the effectiveness of motor response to visual stimuli.
Acknowledgments
The authors would like to thank Mr. Saleem Al Rabadi, Art Director, Vast Technologies, Amman, Jordan, for his professional help in preparing figure 1.
- Received October 7, 2014.
- Accepted May 18, 2015.
- Copyright: © Neurosciences
Neurosciences is an Open Access journal and articles published are distributed under the terms of the Creative Commons Attribution-NonCommercial License (CC BY-NC). Readers may copy, distribute, and display the work for non-commercial purposes with the proper citation of the original work.
In this issue



{kind=link}
Related Articles
Cited By...
- No citing articles found.